La peau est notre plus grand organe et joue un rôle essentiel dans la protection de notre corps. Pourtant, elle est également sujette à un grand nombre de maladies et d’affections qui peuvent affecter sa santé et notre qualité de vie. De l’acné au psoriasis, en passant par l’eczéma ou le mélanome, les maladies de la peau sont nombreuses et souvent mal comprises, malgré leur fréquence croissante.
Les maladies de la peau : Un sujet de santé publique
Les maladies dermatologiques touchent plus de 20 millions de personnes chaque année, et certaines, comme les cancers de la peau, sont en constante augmentation. Beaucoup de ces pathologies, bien qu’elles puissent sembler bénignes, peuvent avoir un impact profond sur le quotidien des malades, tant sur le plan physique qu’émotionnel. La peau est également un reflet de notre bien-être général, ce qui rend ces affections d’autant plus complexes à traiter.
Les principales maladies de la peau
Voici une vue d’ensemble des principales pathologies cutanées :
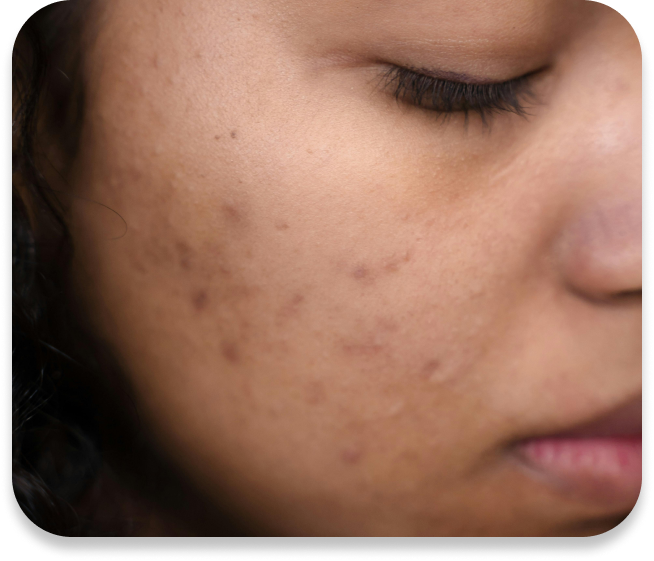
L'acné
Une des affections dermatologiques les plus courantes, en particulier chez les adolescents. Bien que souvent perçue comme bénigne, l’acné peut laisser des cicatrices permanentes et affecter l’estime de soi.
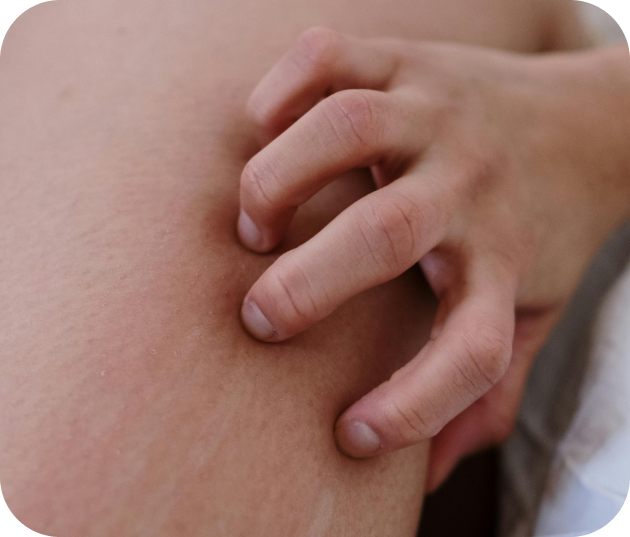
Le psoriasis
Maladie chronique inflammatoire, le psoriasis provoque des plaques rouges et des démangeaisons. Il peut aussi avoir un impact sur la santé articulaire (psoriasis articulaire).
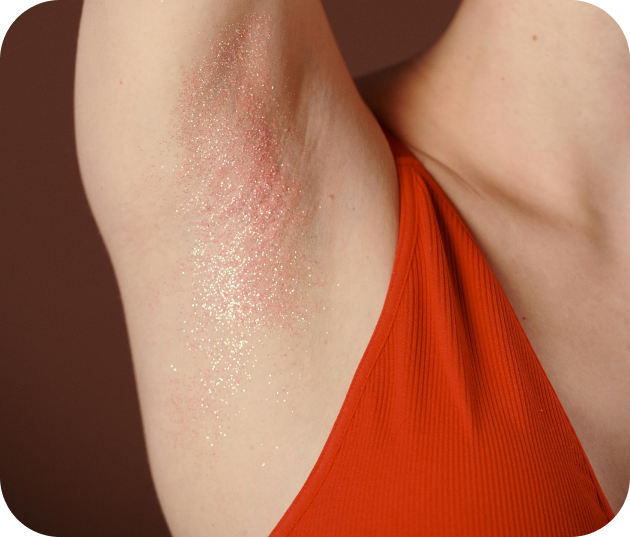
La maladie de Verneuil
La maladie de Verneuil est aujourd’hui décrite comme une maladie inflammatoire chronique du follicule pilosébacé, touchant préférentiellement les zones riches en glandes apocrines (aisselles, plis inguinaux, fesses, sous-mammaire, etc.). »
Les infections cutanées
Elles sont causées par des bactéries, des virus, des champignons ou des parasites. Les infections comme la gale, les impétigos ou l’herpès peuvent être gênantes et parfois très contagieuses.
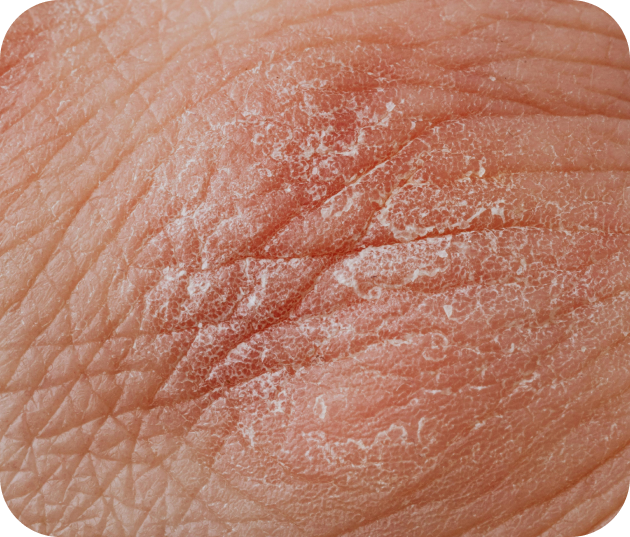
L'eczéma
L’eczéma est une affection inflammatoire de la peau qui provoque des démangeaisons, des rougeurs, une peau sèche et des irritations. Elle touche souvent les enfants, mais peut persister à l’âge adulte.
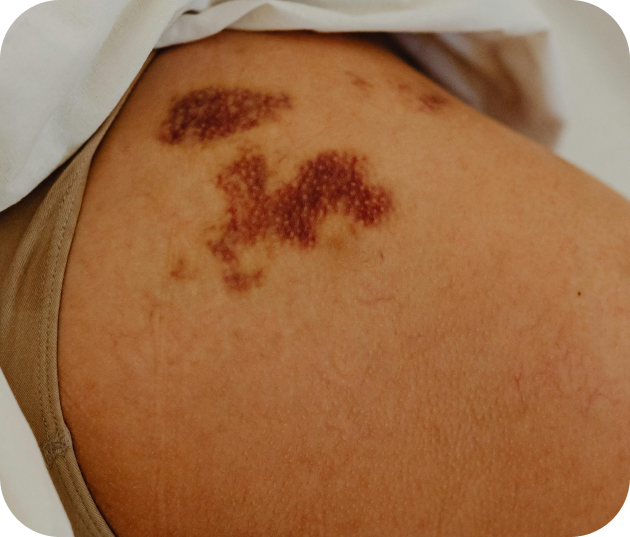
Les cancers de la peau
Le mélanome et les carcinomes basocellulaires ou spinocellulaires sont des cancers cutanés fréquents. La prévention, la détection précoce et un suivi rigoureux sont essentiels pour une prise en charge efficace.
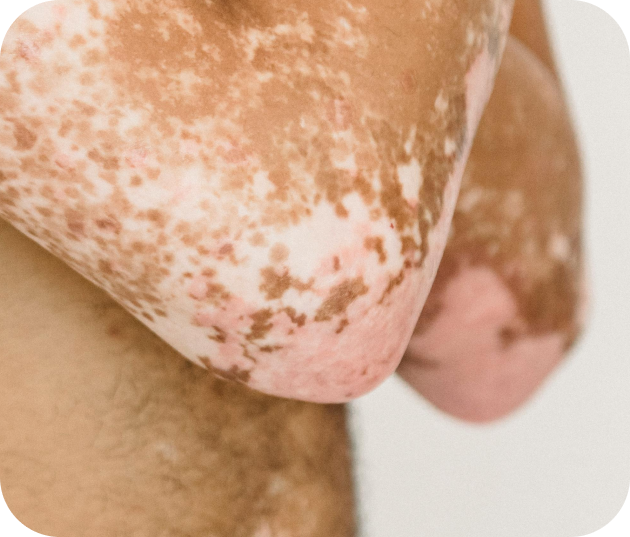
Le vitiligo
Une maladie auto-immune qui provoque des pertes de pigmentation de la peau. Bien que non douloureux, le vitiligo peut avoir un impact psychologique important.
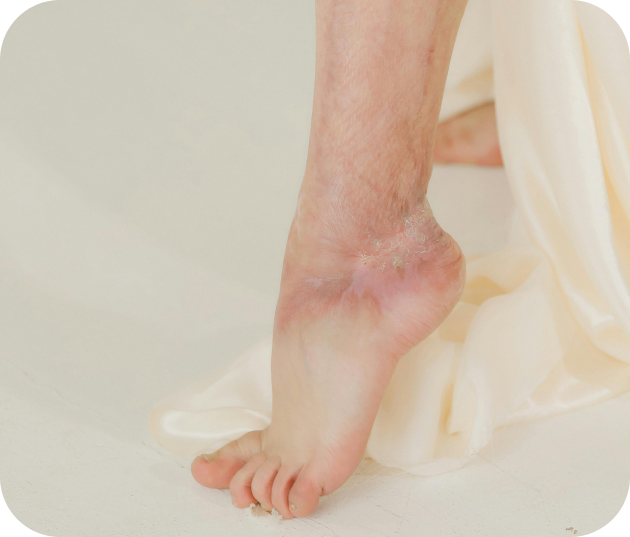
Les maladies rares de la peau
Certaines pathologies dermatologiques sont rares, comme le lupus érythémateux ou les épidermolyses bulleuses, et nécessitent une prise en charge spécialisée.
Les causes des maladies de la peau
Les facteurs génétiques : Certaines maladies de la peau sont héréditaires et peuvent se manifester dès l’enfance.
Les infections : Bactéries, virus, champignons et parasites peuvent causer des infections cutanées.
Les facteurs environnementaux : L’exposition excessive au soleil, la pollution, les produits chimiques et les allergies peuvent aggraver certaines conditions cutanées.
Le stress et l’alimentation : Le stress chronique, une mauvaise alimentation ou certains médicaments peuvent jouer un rôle dans le déclenchement ou l’aggravation des maladies de la peau.
Les maladies systémiques et auto-immunes : Certaines affections cutanées sont liées à des troubles du système immunitaire ou à d’autres maladies générales, comme le diabète ou l’hypothyroïdie.

L'importance de la détection précoce et du traitement
Beaucoup de maladies de la peau peuvent être traitées efficacement lorsqu’elles sont diagnostiquées précocement. Une prise en charge rapide permet d’éviter la progression de la maladie et d’améliorer la qualité de vie des patients. La consultation régulière avec un dermatologue et l’adoption d’un mode de vie adapté sont des éléments clés de la prévention et du traitement.
Prévenir les maladies de la peau
La prévention passe par plusieurs gestes simples mais efficaces :
- Protéger sa peau du soleil : Utiliser des crèmes solaires, porter des vêtements protecteurs et éviter l’exposition excessive.
- Adopter une bonne hygiène : Nettoyer régulièrement sa peau et utiliser des produits adaptés à son type de peau.
- Éviter les facteurs irritants : Certains produits de soin, cosmétiques ou habits peuvent irriter la peau. Il est important de connaître les allergènes et les éviter.
- Consulter un professionnel de santé : En cas de doute ou de symptômes persistants, il est essentiel de consulter un dermatologue pour un diagnostic précis.
- Sensibiliser et informer pour mieux comprendre
Pour en savoir plus sur les maladies de la peau, consultez nos ressources, participez à nos événements et découvrez les initiatives de nos associations membres.